皮膚の特徴 ~皮膚バリア機能とは~
- 監修:
-
- 東京女子医科大学 名誉教授 川島 眞 先生
正常な皮膚と皮膚バリア機能とは
皮膚は体温調節や感覚器としての役割など、様々な機能を持っています。その中でも特に重要な機能として、「皮膚バリア機能」があります。皮膚バリア機能は、角層の内側の水分の蒸散を防ぎ、外界からの異物(アレルゲン・細菌など)の侵入を防御する役割を担っています。
皮膚は外側から表皮、真皮、その下の皮下組織で構成され、表皮はさらに角層、顆粒層、有棘層、基底層から構成されます(図)。
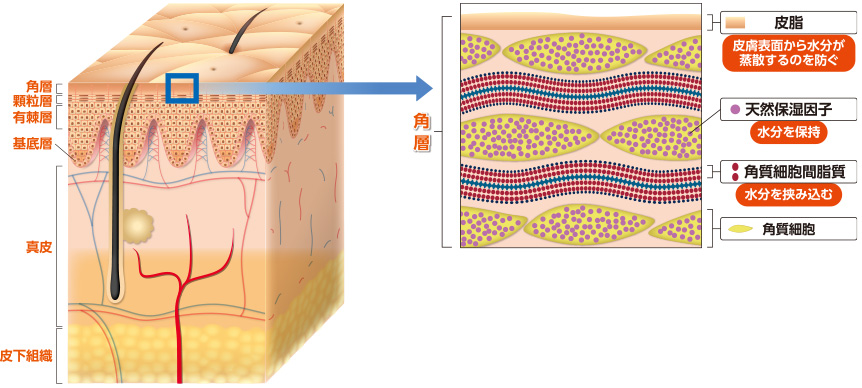
皮膚バリア機能の維持には、皮膚の最外層である「表皮」が最も重要な役割を果たしています。中でも、表皮内の角層は潤いにおける重要な3つの因子、「皮脂」、「角質細胞間脂質」、「天然保湿因子」が存在しています(図)。皮脂は皮脂腺から分泌される脂のことです。汗などと混じり合って皮膚の表面を覆い皮脂膜を形成することで、水分の蒸散を防ぎます。角質細胞間脂質は、角質細胞と角質細胞の隙間を埋めている脂(セラミド、脂肪酸など)です。角質細胞同士を接着させる役割をはたすとともに、水分をサンドイッチ状に挟み込み、逃がさないようにします。天然保湿因子は角質細胞内に存在するアミノ酸や尿素などで、水分を保持します。これらの3つの因子が働き、皮膚の潤いを保つことで皮膚バリア機能を保持しています。
正常な皮膚と乾燥皮膚(ドライスキン)の違い
正常な皮膚(図1)では角層において皮脂、天然保湿因子(アミノ酸や尿素など)、角質細胞間脂質(セラミドや脂肪酸など)が皮膚の潤い(水分量)を一定に保っています。
一方、乾燥した皮膚(ドライスキン:図2)では正常な皮膚と比較してこれらの因子が減少し、角層が構造異常を起こしています。この構造異常により皮膚表面から水分が蒸散しやすく、角層の水分量も減少した状態となっています。また、外界からの異物(アレルゲン・細菌など)が侵入しやすくなっています。このバリア機能の低下を改善するためには、乾燥皮膚を正常な皮膚の状態に近づけることが必要で、そのために保湿剤で潤いを補う必要があります。
図1:正常な皮膚
図2:乾燥皮膚(ドライスキン)
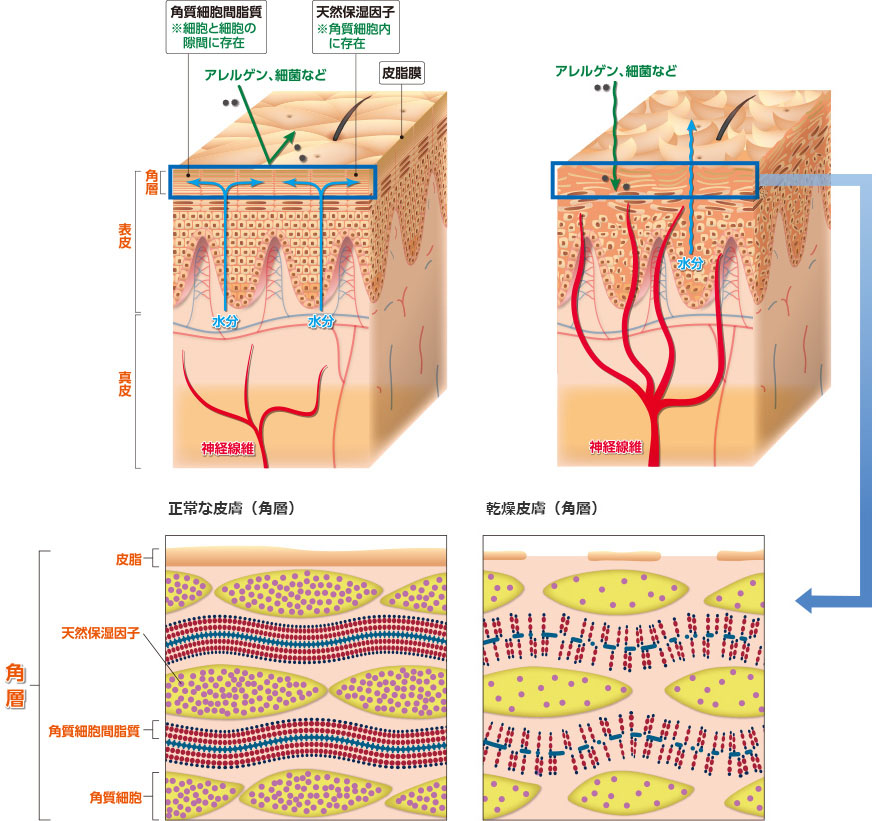
皮膚表面は皮脂で覆われ、角質細胞の隙間を角質細胞間脂質が埋めている。
天然保湿因子も十分に存在し、水分が保持されている。
皮脂、天然保湿因子、角質細胞間脂質の減少により水分保持能が低下して皮膚が乾燥しやすくなる。
乾燥が進むと皮表にひび割れを起こし、皮膚バリア機能が低下する。
乾燥によって痒みが発生するメカニズム
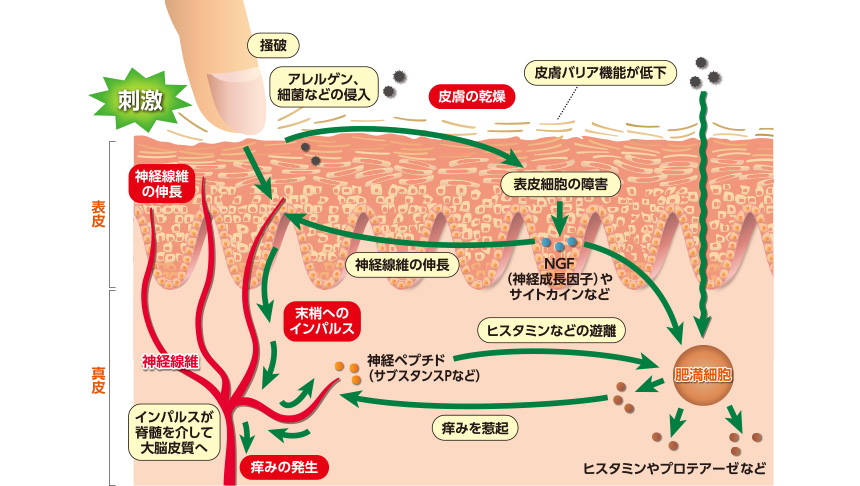
皮膚が乾燥すると、皮膚バリア機能が障害され、同時に物理的または化学的刺激により痒みの神経線維が活性化され表皮内へ伸長します。表皮内に神経線維が増加すると、知覚受容体が増加し、外部からの刺激を感じやすくなり、痒みが発生します。その結果、掻破が誘導され、さらに皮膚表面の角層を破壊して皮膚バリア機能を低下させ、痒みと掻破の悪循環(itch scratch cycle)に陥ります。
また、掻破による外的刺激は表皮細胞を障害し、NGF(nerve growth factor:神経成長因子)やサイトカインなどを産生し、ますます神経線維の伸長を促進します。一方、NGFは真皮内の肥満細胞から、痒みの原因物質であるヒスタミンの放出を誘導します。遊離したヒスタミンは神経線維上のヒスタミン受容体と結合し、痒みのインパルス(電気信号)が中枢と末梢の2方向に伝達されます。中枢へのインパルスは脊髄を介して大脳皮質へと伝達され、痒みが知覚されます。もう一方のインパルスは神経線維の末梢に伝達され、神経ペプチドのサブスタンスPを遊離し、肥満細胞に作用してヒスタミンなどの放出をさらに促進し、痒みを増強させます。
マウスのドライスキンモデルで誘発される表皮内神経伸長に対する保湿剤の効果[参考情報]
乾燥しバリア機能が低下した皮膚では、表皮内神経線維の分布が変化して神経線維の密度が高くなることが知られています。このような皮膚では知覚受容体が増加し、外的刺激により痒みを感じやすくなると考えられるため、乾燥皮膚の早期改善が痒みの抑制に重要です。
最近の研究では、表皮内神経線維の分布は表皮細胞の産生するNGFなどの神経成長因子とSemaphorin3A(Sema3A)などの神経反発因子の量的バランスで制御されていると報告されています1, 2)。しかし、保湿剤がこれらの因子に対してどのような効果を発揮するかは明らかになっていませんでした。そこで、マウスの急性ドライスキンモデルを用いて、ヒルドイドソフト軟膏0.3%が表皮内神経線維の過剰分布や軸索ガイダンス分子であるNGFとSema3Aに与える影響が検討されました。
- Ikoma A et al. : Nat Rev Neurosci, 7(7), 535-547, 2006
- Tominaga M et al. : Br J Dermatol, 158(4), 842-844, 2008
試験概要
-
目的
-
急性ドライスキンモデルマウスを用いて、ヒルドイドソフト軟膏0.3%が表皮内神経線維の過剰分布や軸索ガイダンス分子のNGFとSema3Aに与える影響を検討する。
-
方法
-
雄のICRマウスの皮膚に、アセトンを塗布してドライスキンを作製し、保湿剤の直後塗布群、遅延塗布群、無処置群に分けて検討した。遅延群では、アセトン処理24時間後に保湿剤を塗布、48時間後に表皮内神経線維の分布と数、神経成長因子(NGF)と神経反発因子(Sema3A)の発現量を観察した。対照は無処置群とした。保湿剤はヒルドイドソフト軟膏0.3%を各100mg/Site塗布した。(各群 n=6)
結果
<アセトン処理48時間後>
- 結果① 表皮内神経線維の免疫染色
-
神経線維を免疫染色すると、アセトン処理48時間後では保湿剤を塗布した群の表皮内神経線維の数は、無処置群と比べて減少していた。
記事/インライン画像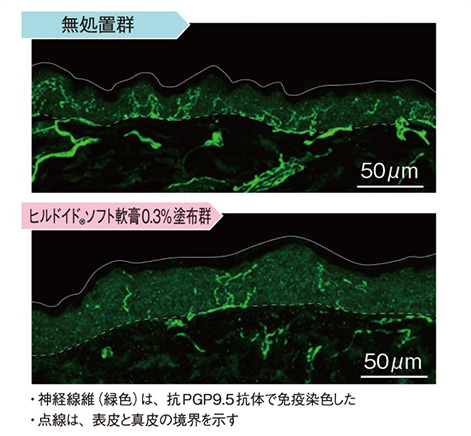
- 結果② 表皮内神経線維数
-
アセトン処理48時間後の表皮内神経線維数は、保湿剤を塗布した群では無処置群と比べて少なかった。
記事/インライン画像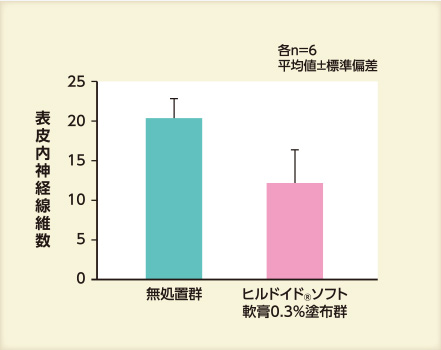
- 結果③ NGF発現量
-
アセトン処理48時間後のNGF発現量は、保湿剤を塗布した群では、無処置群と比べて少なかった。
記事/インライン画像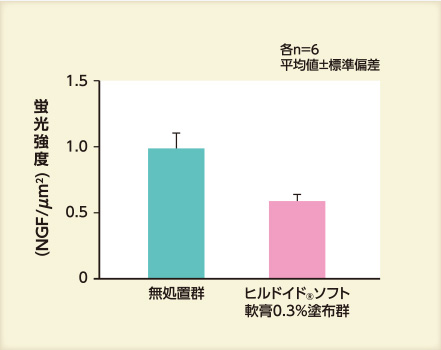
- 結果④ Sema3A
-
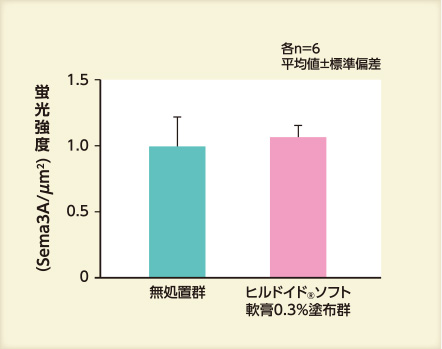
アセトン処理48時間後のいずれにおいてもSema3Aに対する保湿剤の効果は認められなかった。
記事/インライン画像